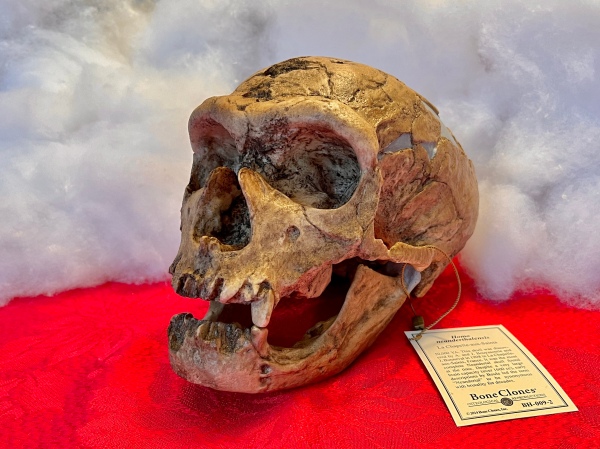
Some years ago, I wrote about the skulls on my desk, asserting that any scientist worth their salt should have at least one. My skulls, however—six of them until recently, are not just “ordinary” modern human skulls (much as I would love to have one), but rather replicas of famous fossil hominid skulls and crania. It has been a while since I’ve added to my collection, but Santa was good to me this past Christmas, bringing me a replica of the “La Chapelle-aux-Saints 1” skull of Homo neanderthalensis, and for today’s birthday my wife gave me a replica of the “Toumaï” cranium of Sahelanthropus tchadensis.
Homo neanderthalensis “La Chapelle-aux-Saints 1”
The “La Chapelle-aux-Saints 1” skull was discovered in 1908 in La Chapelle-aux-Saints, France and is thought to be about 50,000–60,000 years old. It was the most complete Neanderthal skull at the time it was discovered and had a brain capacity exceeding 1600 cc—more than most modern humans. Unfortunately, initial reconstructions of Neanderthal anatomy based on la Chapelle-aux-Saints material depicted the species with thrust-forward skulls, stooped posture, bent hips and knees, and a divergent big toe—reinforcing existing synonymy of the term “Neanderthal” with brutality and savagery. The errors were eventually corrected, but only after decades had passed, and even today this unfair characterization lingers still among the general public.
This particular individual was a male, probably around 40 years of age at the time of his death, and in poor health. He had lost most of his teeth and was suffering from resorption of bone in the mandible and arthritis. This has been widely cited as an example of Neanderthal altruism, since with most of his teeth missing he would have been unable to process his own food. Later studies, however, have shown that the La Chapelle-aux-Saints 1 individual still had enough teeth in place to chew his own food, although perhaps with some difficulty (Tappen 1985).
Sahelanthropus tchadensis “Toumaï”
Sahelanthropus tchadensis was formally described in 2002 based on cranial remains of at least six individuals dated to about 6–7 million years ago during the Miocene epoch. “Toumaï” is the most complete of all the cranial remains, although it was crushed and badly deformed. To date, all the fossils found of Sahelanthropus have come from a small area of northern Chad.
The age of Sahelanthropus puts it around the time of the human-chimpanzee last common ancestor (HCLCA). At the time it was described, only cranial fragments were included in the original description, and the position of the opening for the spinal chord was used to infer that the species walked upright. However, a femur was also found alongside the cranium but was placed with animal bones and excluded from the original analysis. Later analysis of the femur concluded that Sahelanthropus was not bipedal (Macchiarelli et al. 2020), putting its status as a possible relative of the HCLCA into doubt. One alternative possibility that has been raised is that Sahelanthropus is not ancestral to either humans or chimpanzees, but rather to gorillas—a no less significant possibility since fossils attributed to the presumed gorilla lineage at this time consist only of teeth dating to about 10 million years ago.
Tappen, N. C. 1985. The dentition of the “Old Man” of La Chapelle-aux-Saints and inferences concerning Neanderthal behavior. American Journal of Physical Anthropology 67(1):43–50. doi:10.1002/ajpa.1330670106
The pine flatlands of the southeastern Coastal Plain offer an interesting contrast to the upland forests of my home state of Missouri. Closed canopies of oak and hickory are replaced by open canopies of pond and longleaf pine. Dry glades—islands of prairie dotting the forests—are replaced by bogs and bays—oases of wetland punctuating the sandy scrub. In both places, however, fire plays an important role in shaping and preserving these unique habitats. In this ~10,000-acre preserve, prescribed burns spare the heat-tolerant pines—their trunks blackened and scorched but the living branches high above unharmed—and prevent woody shrubs from choking out herbaceous plants, including famously insectivorous plants such as Venus fly traps, sundews, and pitcher plants. I had yet to have seen any of these plants in their native habitats, and after learning of this place and their presence here from a local resident, Madam and I made a beeline to the preserve for an afternoon of botanical hiking.
Pine flatlands at Lewis Ocean Bay Heritage Preserve & Wildlife Management Area.
Almost immediately after starting down the road from the parking lot, I noticed white blossoms dotting the forest floor. Approaching closer revealed them to be wild azaleas—in this case Rhododendron viscosum (swamp azalea). Individually, the petite plants with their crowns of oversized blooms were quaintly beautiful. En masse, clustered on the forest floor, they were an amazing sight to see.
Rhododendron viscosum (swamp azalea) in pine flatland.
Rhododendron viscosum (swamp azalea).
The going was slow in the beginning, with something new to me at every turn. A species of Nuttallanthus (toadflax)—either N. texanus (Texas toadflax) or N. canadensis (Canada toadflax), depending on details of flower dimensions—bloomed abundantly in the sunny openings. A small purple flower was at first assumed to be a species of Tradescantia (spidorwort) but proved to be the related Callisia graminea (grassleaf roseling)—a new genus for me. A bit further down the road we encountered orange flowers that proved to be Polygala lutea (orange milkwort). This genus is represented in Missouri by several species, all having flowers of pink, yellow, or white.
Nuttallanthus (toadflax)—either N. texanus (Texas toadflax) or N. canadensis (Canada toadflax).
Callisia ornata (scrub roseling).
Polygala lutea (orange milkwort).
Flowers were not limited to herbaceous plants. The evergreen woody shrub layer was just coming back to life with new growth, a few of which bore distinctive blossoms identifying them as members of the genus Vaccinium (blueberry). There are several potential species that could be here.
Vaccinium sp. (blueberry).
At one point, Madam called me to the other side of the road, pointing to a strange plant at the edge of a wet area and asking “What’s that?” Bingo—I recognized it instantly as one of the so-called “pitcher plants” (genus Sarracenia), among the most dramatically charismatic of the insectivorous plants. Pitcher plants trap insects using a rolled leaf with downward pointing hairs on the inside and the uppermost part of the leaf flared into a lid (or operculum) to prevent rain from diluting the digestive secretions pooled at the bottom of the leaf. Though a bit past bloom, it was easily identifiable as S. flava (yellow pitcher plant). We were thrilled to have seen our first pitcher plant in the wild, and we looked forward to seeing more (hopefully in full bloom).
Sarracenia flava (yellow pitcher plant).
As we scanned the edge of the wetland looking for more pitcher plants, I noticed tiny white flowers on the small shrubs underfoot. They looked rather “hollyish” to me, and indeed they proved to be Ilex glabra (gallberry), a species of evergreen holly native to the coastal plain of eastern North America and most commonly found in sandy woods and peripheries of swamps and bogs.
Ilex glabra (gallberry).
At this time of season, I was expecting to see insects well active, and this was certainly the case with butterflies—the most common being a species of swallowtail that oxymoronically reminded me of a small giant swallowtail (Papilio cresphontes) but in reality were Papilio palamedes (Palamedes swallowtail). I’m a beetle guy, however, so I was happy to find a few Acmaeodera tubulus jewel beetles on flowers of Erigeron sp. (fleabane), and when I saw a standing recently-dead pine beyond the wet drainage I decided to check it for other jewel beetles on its trunk. As I started to step across the water, a small purplish plant on a piece of wood in the water caught my eye, and I immediately recognized it as one of the sundews (genus Drosera), another genus of insectivorous plants that capture and digest insects using stalked mucilaginous glands that cover their leaf surfaces. This is one of the largest genera of insectivorous plants, but I take this one to be D. intermedia (spoonleaf sundew).
Drosera intermedia (spoonleaf sundew).
Drosera intermedia (spoonleaf sundew).
About a mile and a half from the car, we finally found what we had been hoping to see since soon after we arrived—Sarracenia flava (yellow pitcher plant) in full bloom. A rather large patch of them was visible from afar, their yellow blooms glowing in the sunlight, but sadly most of them were slightly or greatly past peak bloom. A bit further back, however, on the other side of the water, I spotted two single plants in perfect bloom, their petals fresh and intact and making the effort to find a way across the water well worth the effort. Other plants without blooms but with fresh, brightly colored “pitchers” were also seen along the water’s edge.
Sarracenia flava (yellow pitcher plant).
Sarracenia flava (yellow pitcher plant).
Sarracenia flava (yellow pitcher plant).
As we admired the spectacle in front of us, I noticed a clump of red within the vegetation at water’s edge and realized we were looking at another species of pitcher plant—Sarracenia purpurea (purple pitcher plant)! How fortunate we were to see this clump—the only one we saw—which was in perfect bloom and with colorful, freshly-formed pitchers whose squat form contrasted notably from the tall, slender, graceful pitchers of S. flava right next to it.
Sarracenia purpurea (purple pitcher plant).
Sarracenia purpurea (purple pitcher plant).
Sarracenia purpurea (purple pitcher plant).
By now, the April heat had taken a noticeable toll on the conditioning of these two recently-escaped-from-winter Midwesterners. Having found the Holy Grail for the day, we began the long, tired slog back to the car—our legs dragging but our spirits soaring.
Unit tray of Lepturobosca chrysocoma (formerly Cosmosalia chrysocoma).
I belong (or used to belong) to several Facebook groups frequented by insect collectors—both professional and amateur. One question that frequently comes up—primarily for collections with species organized by unit trays—is how to deal with species identification labels. Not surprisingly, the opinions are as varied as the collectors. Some like to put a label on each specimen, while others put a label only on the lead specimen of a series. Some also print or write a separate header label that is placed in the unit tray. What about when names change? Or when reexamination of the specimen(s) reveals an erroneous ID? Should you remove outdated or erroneous identification labels? Fold them? Turn them upside down? Remove them altogether? These questions may seem trivial if one deals only with their own personal collection, but they become much more important when identifying specimens in institutional collections accessible to the public.
Here is my philosophy: an identification is a hypothesis, not data. As a result, ID labels are fundamentally different from labels indicating locality, date, ecological information, and collector, which are data—immutable and unchanging. Identifications can be “wrong” or may change over time, but regardless they merely reflect an individual’s opinion based on their level of expertise and familiarity with state of taxonomy and nomenclature at the time the identification was made. It then follows that identification labels do not need to be placed on every specimen—if a specimen without an ID label gets separated from the series, it does not result in a loss of data in the way it does for a specimen without a locality label, nor do old ID labels need to be changed as a result of nomenclatural changes or corrected identifications—a new label may be added (especially if it is an ID correction), but the old label should not be removed.
Almost as bad as removing old ID labels is folding them, which not only makes them difficult to read but results in mutilation—not just from the fold itself but also from the additional pin holes that are added when re-pinning the folded label. Old ID labels, even if incorrect or out-if-date, represent a historical record of opinion regarding the identity of the specimen, and degrading the labels obscures that history. If one simply must do something to denote a corrected ID, the old label may be turned over, but even then every effort should be made to reuse existing pinholes—just flatten with a fingernail before reusing so the label doesn’t spin. Seriously, however, this simply isn’t necessary—just add the new ID label beneath the old one, which denotes it as the more recent ID (another reason why year should be included on ID labels). Some people don’t like the way this looks, but to do otherwise is to greater priority on visual aesthetics than the integrity of the scientific data represented by the specimen.
As for dealing with nomenclatural changes—I don’t, at least not with already labeled specimens. That old ID label is not “wrong”—it accurately reflects the ID that was given to the series at the time the specimens were identified. Of course, any additional specimens that are added to the unit tray will receive an ID label the reflects the newer nomenclature. Case in point is the above photo, which contains longhorned beetles known for many years as Cosmosalia chrysocoma but recently reassigned to the genus Lepturobosca. You’ll note the older series of specimens bear ID labels with the older name, but the most recently added series contains an ID label with the newer name. There is no reason to go back and change or add ID labels for the older specimens, especially since newer specimens reflecting current nomenclature have been placed in the same unit tray with them. The mix of ID labels representing past and present nomenclature is not problematic—in fact, it adds historical perspective to the series as a whole. On the other hand, were I to receive a series of specimens labeled with an older name from another collection, I would be inclined to add my own, more current ID label (and would certainly do so if the ID—current nomenclature or not—was incorrect), since it was the result of subsequent examination by a different specialist.
Lastly, I don’t waste time creating header labels for unit trays—the ID labels on the specimens themselves are enough to indicate the identity of the species, and the time required to update header labels when nomenclature changes is just that much less time that I have to pin, label, and identify additional specimens being added to the collection.
For the past few years, I’ve been involved with the Missouri Native Plant Society (MONPS). To this point, however, my involvement has been limited to attending the monthly meetings of the St Louis Chapter—unfortunately, now only via Zoom since the beginning of the pandemic. I hope that soon we can return to in-person meetings (or, even better, a hybrid of the two, which allows person-to-person interaction without excluding participation by those who cannot attend in-person), but one activity that has resumed live are their periodic, multi-day field trips. The Spring 2022 Field Trip, held this past weekend in southwestern Missouri, was my first chance to participate in one of these events, and I looked forward to seeing the remnant prairies, limestone, dolomite, and sandstones glades, and chert woodland that were all on tap while rubbing elbows with some of the state’s best botanists and naturalists—some old friends and others new acquaintances!
Day 1 – Schuette Prairie I wasn’t able to make it to the actual Day 1, so I left St. Louis early in the morning to meet the group at the first stop of the following day—Schuette Prairie in Polk Co. Named after my friend and former Cuivre River State Park naturalist, Bruce Schuette, this recently acquired limestone/dolomite prairie with a wet swale contains many plants more typical of glades such as Silphium terebinthinaceum (prairie dock), Echinacea paradoxa (yellow coneflower), and Rudbeckia missouriensis (Missouri coneflower). Of course, on this cold, overcast, early-April morning, it was far too early to see any of these highly charismatic plant species (although some of the more astute botanists were about to point them out by their barely emergent foliage, which was easy to find in the recently-burned northern half of the parcel). Abundantly in bloom, however, was the more subdued Erythronium mesochoreum (prairie fawn lily, midland fawnlily, prairie dogtooth violet). Distinguished from the similar E. albidum (white trout lily) that occurs abundantly further east by its narrower, folded, usually unmottled leaves, all but a few of which remained stubbornly closed against the stiff, cold wind.
Precious few other blooms were seen—I recall somebody mentioning they had seen Viola sororia (common violet), and I photographed this little clump of Fragaria virginica (wild strawberry) that will eventually provide food for one of the area’s many box turtles.
Fragaria virginica (wild strawberry).
Speaking of box turtles, I found this completely naked, bleached carapace and at first hoped that it might have been from an ornate box turtle (Terrapene ornata)—limited in Missouri to western prairies and a species I have not yet seen. However, the presence of a midline ridge and its relatively more domed shape suggest it is from a three-toed box turtle (Terrapene carolina triunguis).
Many other carapaces were seen (though none in such good shape), and in fact bones of many types were easy to find in the burned portion of the prairie. This disarticulated skull from what appears to be a young calf (Bos taurus) was perhaps the most impressive bone find, but we did also find a dried skeleton of a smaller individual. Being the lone entomologist of the group, I just had to turn over the carcass and search for beetles and managed to capture a skin beetle (family Trogidae) and one other small unidentified beetle (but, unfortunately, no Necrobia rufipes [red-legged ham beetle]).
Disarticulated bovid skull – probably a young calf (Bos taurus).
Rocky Barrens Conservation Area Later in the morning, the group caravaned to Rocky Barrens Conservation Area, a 281-acre area in Greene Co. featuring Mississippian limestone glades and site for the federally-endangered Physaria filiformis (Missouri bladder-pod). This plant, in the mustard family, is found only in four counties in southwest Missouri. The plants were readily found, but we were too early to see them in bloom—or anything else, for that matter. For me, however, the glade alone was still interesting, and I couldn’t help but take note of the similarities—and differences—between this limestone example and the dolomite glades south of St. Louis with which I am so much more familiar. Almost immediately, I noted the presence of Sideroxylon lanuginosum (gum bumelia), host for Plinthocoelium suaveolens (bumelia borer)—surely one of North America’s most beautiful longhorned beetles! I didn’t see any frass piles at the base of any of the trees, the presence of which would indicate larval activity, but I’m sure the beetle is here. It would be interesting to come back during the season and look for it. While I didn’t find any signs of the beetle, I couldn’t miss the bright orange-yellow gold-eye lichens (Teloschistes chrysophthalmus) colonizing it’s branches.
Teloschistes chrysophthalmus (gold-eye lichen) on branch of living Sideroxylon lanuginosum (gum bumelia).
Another tree that caught my interest was Celtis tenuifolia (dwarf hackberry). I see these small, gnarly versions of the genus in glades and other xeric habitats, and they always catch my interest because of the diversity of interesting woodboring beetles associated with it. As I looked at the trees, I noticed one small tree in particular that was the perfect stage of dead—branches brittle but bark mostly still intact with a little bit of peeling on the trunk revealing woodboring beetle larval galleries underneath! There were only a few emergence holes present—strong evidence that the tree was still infested and worth bringing back to put in an emergence box to trap the emerging adult beetles. With luck, I’ll be pinning a series of Agrilus ferrisi next winter!
Corry Flatrocks Conservation Area After lunch at a nearby city park, the group caravaned to Corry Flatrocks Consevation Area in Dade Co.—site of another federally-endangered plant, Mononeuria minima (formerly Geocarpon minima) (tiny-Tim, earth fruit). The sandstone glades at this site are among the largest in the area and, thus, host a large population of the plant. By this time of day, the sun had been out for awhile and the day had warmed considerably, so we hoped to see other flowering plants as well. Among the first that we encountered while walking towards the glade proper was Ranunculus fascicularis (early buttercup), distinguished from other “large-flowered buttercups” by its canescent (grayish due to hairiness) leaves with long and narrow lobes, their tips bluntly pointed or rounded. The dry, gladey habitat also distinguishes the species from the similar R. hispidus (hairy buttercup), which flowers at the same time but prefers moister habitats.
Ranunculus fascicularis (early buttercup).
On the glade proper, we quickly encountered tiny little saxifrages in bloom, which turned out to be Micranthes texana (Texas saxifrage), restricted in Missouri to this part of the state (and thus with a high CC value of 9) and distinguished from the more widespread M. virginiensis (early saxifrage) by its small, compact stature. These first individuals we encountered had especially reddish-tinged flowers.
Micranthes texana (Texas saxifrage).
Micranthes texana (Texas saxifrage).
As soon as we reached the more open part of the glade with large expanses Of exposed rock, the group dropped to their hands and knees to find the diminutive plants we were looking for.
MONPS Field Trip participants looking for Mononeuria minima (formerly Geocarpon minima) (tiny-Tim, earth fruit).
The plants were not uncommon, even abundant, in shallow, sand-filled depressions in the rock. Nevertheless, careful observation was still required to see and recognize them. Fortunately, the plants were already in bloom, their tiny styles barely visible to the naked eye within the green, not-much-bigger, petalless flowers. Photographing these plants, and especially those in bloom, proved to be a task almost beyond the capabilities of the smart phones that most in the group were using (me included).
Mononeuria minima (formerly Geocarpon minima) (tiny-Tim, earth fruit).
Mononeuria minima (formerly Geocarpon minima) (tiny-Tim, earth fruit).
The glades stretched on for quite a distance, inviting further exploration. At the margins, white flowering trees were noticed, and moving closer they proved to be Amelanchier arborea (downy serviceberry, common serviceberry)—among the first we have seen open this spring. (I typically see the first blooms of these trees in the final days of March, at least around my home in east-central Missouri.) an even closer looked revealed tiny insects (also among the first insects I have seen active this spring) flying around and crawling about on the flowers. These proved to be parasitic hymenopterans—family ID is still pending, but I suspect they will prove to be a species in one of the many families of “microhymenopterans” that are egg parasitoids. I am not sure whether they were visiting the flowers as pollinators (which behavior I am not aware of) or in hopes of encountering other pollinators which could potentially serve as hosts—a subject with which I will need to follow up.
Amelanchier arborea (downy serviceberry, common serviceberry) with numerous tiny parasitic wasps (family undetermined).
Near the back end of the glade, we encountered a few more Micranthes texana (Texas saxifrage), these having more typical white flowers in perfect peak bloom.
Micranthes texana (Texas saxifrage) with white flowers.
Micranthes texana (Texas saxifrage) with white flowers.
Also in that part of the glade we found a few scattered individuals of Selenia aurea (golden selenia). While not quite as conservative as M. texana (CC value = 6), it has a similar range in the U.S. and in Missouri is also restricted to a handful of counties in the southwestern part of the state. The plant is known to occur in large colonies (which I have seen at nearby Corry Branch Glade)—its brilliant yellow flowers forming a spectacular display.
Selenia aurea (golden selenia).
To this point, the only insect I had seen besides the microhymenopterans was a skin beetle (family Trogidae), which I found when I kicked over some dried mammal scats. However, on the way back to the cars we finally encountered an insect large enough in size and striking enough in appearance to pique the interest of not just me but the group as a whole—a large caterpillar feeding on the foliage of Penstemon digitalis (smooth beard-tongue). It’s appearance—dark with longitudinal yellow stripes and blue spotting—immediately called to mind one of the tiger moths (formerly Arctiidae, now a subfamily in the Erebidae), specifically the genus Haploa (commonly called haploa moths). A little detective work on BugGuide comparing photos and recorded host plants narrowed the likely choice to H. confusa (confused haploa moth).
Haploa sp. prob. confusa (confused haploa moth) caterpillar feeding on foliage of Penstemon digitalis (smooth beard-tongue).
Day 2 – Lead Mines Conservation Area The final day of the MONPS Field Trip featured a morning trip to Lead Mine Conservation Area in Dallas Co. Of particular interest to the group were several parcels within the area designated as Niangua River Hills Natural Area and featuring a diversity of habitats including dolomite glades, chert woodlands, and calcareous wet meadows (fens). Most in the group visited the northern parcel to see the dolomite glades; however, a few of us—primarily from St. Louis and well-familiar with dolomite glades—opted to visit the smaller southern unit of the natural area to see the fen and riparian woodland we needs to pass through to get there. It was a much warmer morning than yesterday, though still chilly starting out, so blooms were sparse as we hiked the woodland trail searching for any hint of color. At one point, someone noticed a shrub a bit off the trail with large, reddish pink flowers—the color seeming a bit unexpected for the situation. Bushwhacking toward it, we realized it was Chaenomelesspeciosa (common flowering quince), a common, ornamental non-native plant that rarely—but obviously sometimes—escapes cultivation. While the group looked at the plant, I saw my first insect of the day—Paraulacizes irrorata (speckled sharpshooter), one of our largest and most recognizable leafhoppers, sitting head-down on the stem of a small sapling.
Paraulacizes irrorata (speckled sharpshooter).
Among the first native blooms we saw was Ranunculus hispidus (hairy buttercup). Though similarly “large-flowered” as R. fascicularis (early buttercup), it differs by its sprawling growth habit, differently shaped-leaves, and preference for moist habitats. Buttercups are a favorite flower host for jewel beetles (family Buprestidae) in the genus Acmaeodera, and one species —A. tubulus—is among our earliest-emerging beetles in the spring, so I checked each buttercup flower that I saw hoping to see these little beetles signaling the beginning of insect activity for the season. Sadly, none were seen.
Ranunculus hispidus (hairy buttercup).
At last we reached the fen—a large open area on the toe-slopes of the adjacent hillside where water draining through the underlying strata emerged to the surface to maintain a continually wet environment. The fen here is special, as two species of Cyprepedium (lady’s slipper orchids) are know to occur in the fen (and in fact, all four of the state’s Cyprepedium spp. can be found with Lead Mine Conservation Area). At this early date, the orchids would not be anywhere close to blooming; however, the group looked for evidence of their presence, walking gingerly through the fen so as to avoid inadvertently stepping upon any emergent foliage. No putative clumps were found, but already in my mind I’m thinking a mid-May trip back to the fen might be warranted! Unlike the orchids, Castilleja coccinea (Indian paintbrush) was abundantly evident throughout the fen, with an occasional plant almost ready to burst forth their scarlet blooms. Senescent flower stems of composites, presumably Rudbeckia, were also seen throughout the glade, which, combined with the abundance of Castilleja, created the promise of a stunning early-summer display across the fen.
Castilleja coccinea (Indian paintbrush).
During our time in the fen, two species of butterflies were seen flitting about the herbaceous vegetation: tiny blue Celastrina ladon (spring azure), and one of the dustywing skippers in the genus Erynnis. The former were impossible to photograph due to their persistent flitting and skittish behavior, and the latter almost were as well. Only when I locked the focus on a preset 2x zoom and fired shots in rapid succession while moving the smartphone ever closer to the subject did I manage this one imperfect but passable photograph of the last one I tried. The genus Erynnis is diverse and notoriously difficult to identify, and my expertise with skippers and butterflies pales compared to my skills with beetles, so the ID will have to remain Erynnis sp. until a more authoritative opinion is offered. [Edit 4/6/22, 11:38 am: According to my lepidopterist friend Phillip Koenig, Erynnis horatius and E. juvenalis both fly in early spring, and they cannot be reliably separated from the dorsal side. Erynnis juvenalis has one or two dots on the ventral hind wing that E. horatius lacks and only flies in the early spring, while E. horatius can be seen through the summer. If only I could turn the picture over to see what it looks like on the ventral side!]
Erynnis horatio or E. juvenalis (Horatio’s or Junenale’s duskywing) in fen habitat.
Returning through the riparian woodlands after visiting the fen, the day had warmed considerably, and numerous flowers not seen earlier were suddenly in full bloom. These included Erythronium mesochorium (prairie fawn lily midland fawnlily, prairie dogtooth violet)—the same species we saw yesterday so reluctantly in bloom at Schuette Prairie. Most were of the familiar form with unmottled leaves; however, we found one individual with notably mottled leaves that resembled those of E. albidum (white dogtooth violet) (1st photo). Nevertheless, the leaves were still narrower than that species and folded, and the plant was growing a mere 12” from another individual with no trace of mottling (2nd photo).
Claytonia virginca (spring beauty) was also blooming in abundance as we took the trail back. I am always amazed at the variability seen in the flowers of this species—from pure white to vividly pink-striped to pink at the tips. This especially vivid pink individual was about as pink as they come.
Claytonia virginca (spring beauty)—an especially vivid pink example.
Sanguinaria canadensis (bloodroot) also was popping up regularly. We had seen isolated plants sitting the trailsides when we first part through—their flowers tightly folded in stubborn response to the chilly morning temperatures. By early afternoon, however, they were spread wide open as invitation to any of the flying insects that had surely also been awakened by the warmer temperatures of the afternoon. While most were seen as isolated individuals, a particularly idyllic clump captured our attention, almost begging “photograph me!”
Sanguinaria canadensis (bloodroot).
Sanguinaria canadensis (bloodroot).
With that, we rejoined the main group to recount the days experiences and cement new relationships before heading back towards our respective home areas.
Long Ridge Conservation Area On the way back home, I decided to check out this conservation area in Franklin Co., which I’ve never visited before. The afternoon had gotten quite warm, so I reasoned that maybe today would be the day when insects start coming out in abundance. I was right! As soon as I pulled into the parking lot, I saw a Prunus mexicana (Mexican plum) in full bloom, and walking up to it I immediately saw an abundance of bees and small beetles all over the flowers. The latter turned out to be Orsodacne atra (a leaf beetle) and Ischnomera ruficollis (rednecked false blister beetles).
Orsodacne atra (leaf beetle) on flowers of Prunus mexicana (Mexican plum).
Orsodacne atra (leaf beetle) mating pair on flowers of Prunus mexicana (Mexican plum).
Ischnomera ruficollis (rednecked false blister beetle) mating pair on flowers of Prunus mexicana (Mexican plum).
Inside the woods along the Blue Trail, there were the usual suspects in bloom—Claytonia virginica (spring beauty), Cardamine concatenata (toothwort), Antennaria parlinii (Parlin’s pussytoes) and Ranunculus hispidus (hairy buttercups).
Antennaria parlinii (Parlin’s pussytoes).
Eventually I happened upon an Amelanchier arborea (downy serviceberry) in full bloom. There were more O. atra and I. ruficollis on the flowers (though not so many as on the Mexican plum), along with a Mecaphesa sp. crab spider that had caught and was feeding on a male Andrena carlini (Carlin’s mining bee)*.
Mecaphesa sp. crab spider with male Andrena carlini (Carlin’s mining bee) prey on flowers of Amelanchier arborea (downy serviceberry). *Bee ID by Mike Arduser.
On the back third of the trail, I found two fallen branches under a Quercus shumardii (Shumard’s oak) that had been pruned by longhorned beetles—presumably Anelaphus villosus. At the end of the trail I found a third such branch of the same species of oak. All three will be placed in an emergence box, and hopefully the culprits will emerge as adults.
Anelaphus villosus-pruned branches of Quercus shumardii (Shumard’s oak)—both collected under the same tree.